2.2 Způsoby získávání energie
2.2.1. Anaerobní způsob získávání energie
Je charakterizován možností svalových buněk vykonávat mechanickou práci při využívání energie uvolněné bez účasti kyslíku. Anaerobní zdroje energie využívá organismus v situacích, kdy není schopen zabezpečit dostatek energie efektivnějším aerobním způsobem.
Aktuální potřeba energie přesahuje rychlost mobilizace aerobních procesů (daných především funkcí transportního systému) na začátku zátěže, při náhlém zvýšení intenzity svalové práce nebo při vysoké intenzitě svalové práce po překročení maximálního množství kyslíku, které je systém schopný využít (Meško, 2005).
Podle převažujícího zdroje energie se anaerobní systém získávání energie dělí na způsob anaerobní alaktátový – energie je uvolněna z ATP a CP (kreatinfosfát) bez účasti anaerobní glykolýzy a tvorby laktátu (ATP-CP systém) a způsob anaerobně laktátový, kdy je energie získána z anaerobní glykolýzy s tvorbou laktátu.
Biochemické reakce:
ATP → ADP + energie pro svalový stah
CP + ADP → ATP + C
Glukóza → ATP + 2 LA
2.2.2. Aerobní způsob získávání energie
Tento způsob získávání ATP je dominantní při tělesných aktivitách vytrvalostního charakteru trvajícího déle než 2–3 minuty (Meško, 2005). Úroveň aerobních schopností je ovlivněna dědičností (80%). Aerobní schopnosti jsou limitujícím faktorem výkonnosti ve vytrvalostních disciplínách a o její úrovni nás informuje vrcholová spotřeba kyslíku (VO2,peak) – maximální množství kyslíku přijaté organismem při zátěžovém testu se zátěží do subjektivního maxima spojeném s analýzou vydechovaných plynů při spiroergometrii.
Biochemická reakce:
glukóza + 6 O2 → 36 ATP + 6 H2O + 6 CO2
Obrázek 2.2. Podíl zdrojů energie na její celkové úhradě v závislosti na čase při maximálních výkonech různého trvání
 Rychlostní zatížení s dobou trvání výkonu přibližně 15 s využívá jako hlavní energetický zdroj systém makroergních (na energii bohatých) fosfátů ATP a CP (ATP – CP systém) s nepatrnou tvorbou laktátu. Rychlostně vytrvalostní zatížení od 15 – 50 s využívá ATP a CP, navíc anaerobní glykolýzu s tvorbou laktátu. Zdrojem energie při vytrvalostním krátkodobém zatížení do 2 min je anaerobní glykolýza s velmi vysokou tvorbou laktátu (glykolytická fosforylace). Vytrvalostní zatížení střední 2 – 11 min využívá především glycidy se střední tvorbou laktátu, dlouhé vytrvalostní zatížení 11 – 60 min využívá glycidy a lipidy, tvorba laktátu je malá. Velmi dlouhá doba zatížení delší než 60 min využívá jako energetický zdroj převážně lipidy a glycidy, laktát se tvoří v malé míře (Havlíčková, 2004).
Rychlostní zatížení s dobou trvání výkonu přibližně 15 s využívá jako hlavní energetický zdroj systém makroergních (na energii bohatých) fosfátů ATP a CP (ATP – CP systém) s nepatrnou tvorbou laktátu. Rychlostně vytrvalostní zatížení od 15 – 50 s využívá ATP a CP, navíc anaerobní glykolýzu s tvorbou laktátu. Zdrojem energie při vytrvalostním krátkodobém zatížení do 2 min je anaerobní glykolýza s velmi vysokou tvorbou laktátu (glykolytická fosforylace). Vytrvalostní zatížení střední 2 – 11 min využívá především glycidy se střední tvorbou laktátu, dlouhé vytrvalostní zatížení 11 – 60 min využívá glycidy a lipidy, tvorba laktátu je malá. Velmi dlouhá doba zatížení delší než 60 min využívá jako energetický zdroj převážně lipidy a glycidy, laktát se tvoří v malé míře (Havlíčková, 2004).
2.2.3. Kyslíkový deficit a kyslíkový dluh
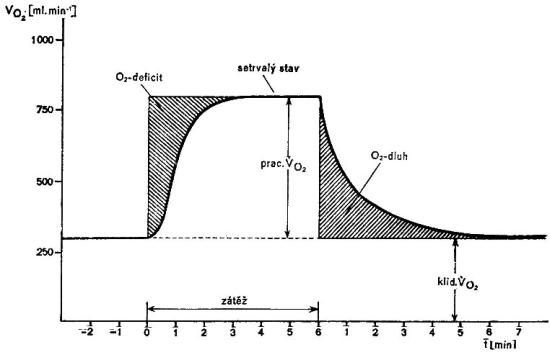
Bezprostředně po zahájení práce není schopen transportní systém dodat dostatečné množství kyslíku pracujícím tkáním. Existuje nepoměr mezi jeho nabídkou a poptávkou, rozvíjí se kyslíkový deficit, který se splácí po ukončení zátěže formou kyslíkového dluhu. Kyslíkový dluh (pozátěžový kyslík) představuje veškerou nadspotřebu kyslíku nad klidovou hodnotu po ukončení zátěže a je považován za kvantitativní měřítko anaerobního metabolismu. Skládá se ze 3 složek. První rychlá alaktátová slouží k obnově ATP a CP během prvních 2 – 3 minut po ukončení zátěže. Druhá složka je pomalá laktátová a vede k resyntéze glykogenu, třetí pomalá alaktátová obnovuje „dolaďuje“ klidové funkčně metabolické podmínky.
Obrázek 2.3. Kyslíkový dluh a kyslíkový deficit
 2.2.4. Anaerobní práh
2.2.4. Anaerobní práh
Je taková intenzita zátěže (rychlost běhu, výkon na ergometru), kdy se k převážně aerobnímu způsobu krytí energetických požadavků organismu přidává a dále narůstá způsob anaerobní. Důsledkem je zvýšená tvorba a kumulace kyseliny mléčné se zvýšením koncentrace vodíkových iontů. Stálá hladina vodíkových iontů je nezbytnou podmínkou, nutnou k funkci buněk organismu. Proto dochází k aktivaci kompenzačních mechanismů, které udržují stálost vnitřního prostředí (homeostázu). Nejdůležitější je nárazníkový bikarbonátový systém.
Slabá kyselina uhličitá disociuje na vodíkový iont a hydrouhličitan sodný (bikarbonát), současně je však v dynamické rovnováze s rozpuštěným oxidem uhličitým:
H+ + HCO3− == CO2 + H2O
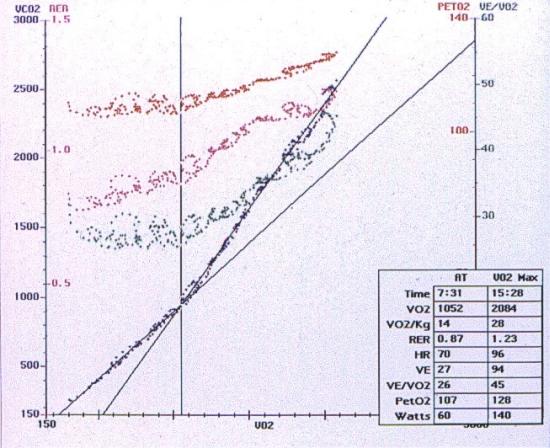
Složky systému jsou bezprostředně regulovatelné prostřednictvím respiračního systému. Zvýšení hladiny vodíkových iontů vede k jejich vyššímu slučování s bikarbonátem, a tím k jeho poklesu. Zvyšuje se produkce CO2 (rovnice se „posunuje doprava“), dochází k podráždění dýchacího centra, a tím k hyperventilaci, při které je zvýšeně produkovaný oxid uhličitý z organismu eliminován. Stanovení anaerobního prahu lze provést buď neinvazivně (bez zásahu do organismu) z ventilačně respiračních hodnot získaných při spiroergometrickém vyšetření jako ventilační práh (ventilatory threshold – VT), nebo invazivně z hodnot laktátu či úbytku bází (bikarbonátů) získaných odběrem krve při zátěži. Anaerobní práh je určen přechodem k strmějšímu vzrůstu („zlomem křivky“) vývoje sledovaných hodnot.
Obrázek 2.4. Příklad určení „ventilačního“ prahu při spiroergometrickém vyšetření z křivky vývoje výdeje CO2 a ostatních respiračně ventilačních parametrů
 Při vytrvalostním tréninku by intenzita cvičení měla vzhledem k hodnotě VO2max být co nejvyšší, ale neměla by vést k výraznější produkci kyseliny mléčné, a tím k ovlivnění vnitřního prostředí. Těmto požadavkům odpovídá intenzita zátěže blízká úrovni anaerobního prahu jako nejvyšší možná úroveň pracovní činnosti, při níž k úhradě energie slouží především aerobní procesy. Produkce kyseliny mléčné odpovídá její spotřebě. Z těchto důvodů je také úroveň anaerobního prahu považována za relativně bezpečný limit intenzity zátěže i při doporučení fyzické aktivity nemocným. Při překročení ANP by mohlo dojít k prudkému zvýšení koncentrace vodíkových iontů (rozvoji metabolické acidózy) a poškození zdraví.
Při vytrvalostním tréninku by intenzita cvičení měla vzhledem k hodnotě VO2max být co nejvyšší, ale neměla by vést k výraznější produkci kyseliny mléčné, a tím k ovlivnění vnitřního prostředí. Těmto požadavkům odpovídá intenzita zátěže blízká úrovni anaerobního prahu jako nejvyšší možná úroveň pracovní činnosti, při níž k úhradě energie slouží především aerobní procesy. Produkce kyseliny mléčné odpovídá její spotřebě. Z těchto důvodů je také úroveň anaerobního prahu považována za relativně bezpečný limit intenzity zátěže i při doporučení fyzické aktivity nemocným. Při překročení ANP by mohlo dojít k prudkému zvýšení koncentrace vodíkových iontů (rozvoji metabolické acidózy) a poškození zdraví.